ウイルス分離状況から見た2002/03シーズンのインフルエンザの流行の特徴は、 1)流行の開始時期が前シーズンより約1カ月早く、 そのピークは第2~6週目であったこと、 2)ここ2シーズンはA/H1型、 A/H3型、 B型ウイルスの混合流行であったが当該シーズンはA/H1型の流行はなく、 A/H3型とB型(2:1)の混合流行であったことがあげられる。
1.ウイルス抗原解析
全国の地方衛生研究所(地研)で分離されたウイルス株は、 国立感染症研究所(感染研)からシーズン前に配布された抗原解析用抗体キット[A/New Caledonia/20/99 (A/NC/99, H1N1)、 A/Moscow/13/98(H1N1)、 A/Panama/2007/99 (A/PA/99, H3N2)、 B/Hiroshima(広島)/23/2001(山形系統)、 B/Shandong(山東)/7/97(Victoria系統)に対するフェレット感染血清およびB/Kagoshima(鹿児島)/11/2002(Victoria系統の主流行株、 高度免疫羊血清)]を用いた赤血球凝集抑制(HI)試験で、 各地研において型別同定および抗原解析が行われた。感染研ではこれらの成績をもとにして、 HI価の違いの比率が反映されるように選択した分離株(分離総数の約5%に相当する)について、 A/H1N1ウイルスには5種類、 A/H3N2ウイルスおよびB型ウイルスにはそれぞれ9種類の参照抗血清を用いてさらに詳細な抗原解析を行った。
1)A/H1N1ウイルス:2002/03シーズンではH1N1ウイルスは1株のみ分離され、 その抗原性はワクチン株A/NC/99に類似していた(表1)。
一方、 諸外国においては、 ヨーロッパ、 米国でH1ウイルスの流行があったが、 いずれの地域においてもワクチン株A/NC/99類似株が主流で、 特別な変異株の出現は観察されなかった。また、 2001/02シーズンに初めて捉えられた遺伝子再集合体H1N2ウイルス(WHO, WER, 77, 77-80, 2002および本月報Vol.23, 122, 2002参照)も引き続き分離されたが、 その抗原性は前シーズン分離株と同じであった。
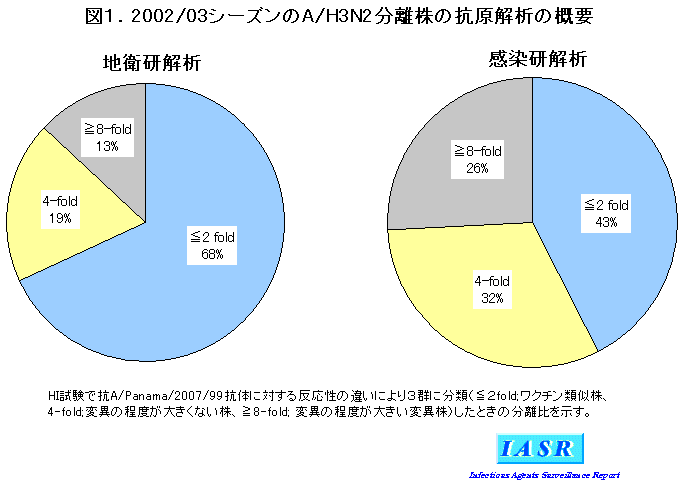
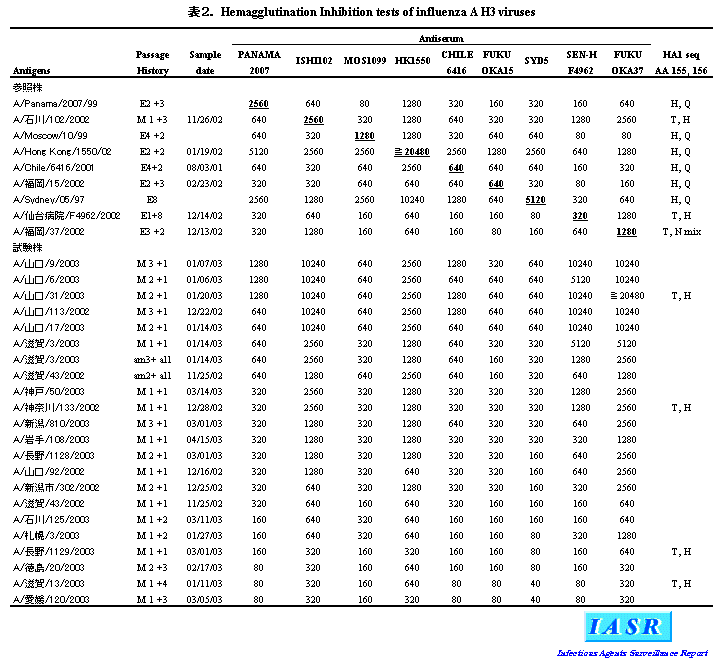
2)A/H3N2ウイルス:地研での解析では、 分離株の大半は2000/01シーズンから採用されているワクチン株A/PA/99類似株であったが、 HI試験でA/PA/99から4倍以上の抗原変異をした株も32%と前シーズンより増えてきた(図1)。感染研での詳細な抗原解析では、 変異株の占める割合が58%であった(図1)。分離株のHI試験の成績の一部を表2に示したが、 これら変異株はその代表株であるA/Ishikawa(石川)/102/ 2002やA/Sendai-H(仙台病院)/F4962/2002 抗血清とよく反応した。また、 変異株の多くはHA蛋白の抗原決定領域Bに入る155、 156番目のアミノ酸に特徴的な置換(155Tおよび156H)をもっており、 系統樹解析においても前シーズン分離株とは区別できる1群を形成していた(図3)。さらに、 諸外国での分析においても同様の結果が得られていることから、 WHOは2004シーズンの南半球のインフルエンザワクチンとしてA/Fujian(福建)/411/2002類似株[ワクチン製造株としては孵化鶏卵で高増殖するA/Kumamoto(熊本)/102/2002 またはA/Wyoming/3/2003]を推奨した(WHO, WER, 78, 375-379, 2003参照)。
3)B型ウイルス:B型インフルエンザウイルスには、 B/Yamagata(山形)/16/88で代表される山形系統とB/Victoria/2/87で代表されるVictoria系統がある。2002/03シーズンは前シーズンに引き続きVictoria系統株がわが国をはじめ世界の主流であった(国内での分離比はVictoria系統:山形系統=97:3)。
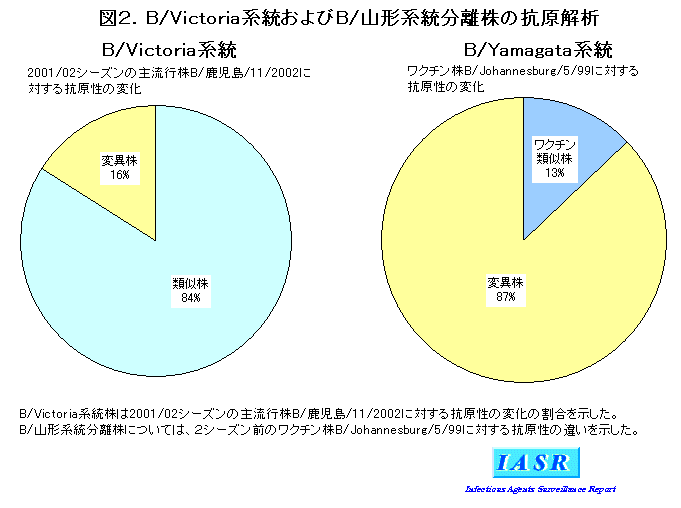
表3にB型分離株のHI試験の成績の一部を示す。Victoria系統に入るMDCK細胞分離株の多くは、 孵化鶏卵で増殖させたウイルスを抗原として作製したフェレット抗血清(抗B/Shandong/7/97 抗血清や抗B/Hong Kong(香港)/330/2001抗血清)に対して低いHI価しか示さないことから、 MDCK細胞を用いて分離された流行株については、 標準株からの抗原性のズレの程度や流行株間での抗原性の多様性などは特定できない(本月報Vol.23、 279-287、 2002参照)。そこで、 当該シーズンには、 前シーズンの流行の主流であった分離株の中からMDCK細胞でよく増えるB/Kagoshima/11/2002 株を選定し、 これを抗原とした高度免疫羊血清を作製し、 各地研に配布した。抗B/Kagoshima/11/2002血清は高度免疫血清ではあるが、 抗原性の変化を比較的明確に捉えることができた。この結果、 分離株の多くは抗B/Kagoshima/11/2002血清によく反応したが、 HI試験で8倍以上の違いを示す変異株も全体の16%を占めていた(図2、 表3)。このことは、 当該シーズンの主流であったVictoria系統株の流行は、 前シーズンと同じ傾向であったことを示している。
一方、 少数ながら分離された山形系統株の多くは、 2シーズン前のワクチン株B/Johannesburg/5/99からは抗原性が大きく変化していた(図2、 表3)。このことは、 山形系統株が再び流行の主流になる場合に備えて、 変異株群からワクチン候補株となる孵化鶏卵分離株を準備しておく必要があることを示している。
2.ウイルスHA遺伝子の解析
1)A/H1N1ウイルス:2002/03シーズンはワクチン類似株が1株しか分離されなかったことや、 世界的にも大きな変化は見られなかったことから、 A/H1ウイルスの系統樹の掲載は割愛する。
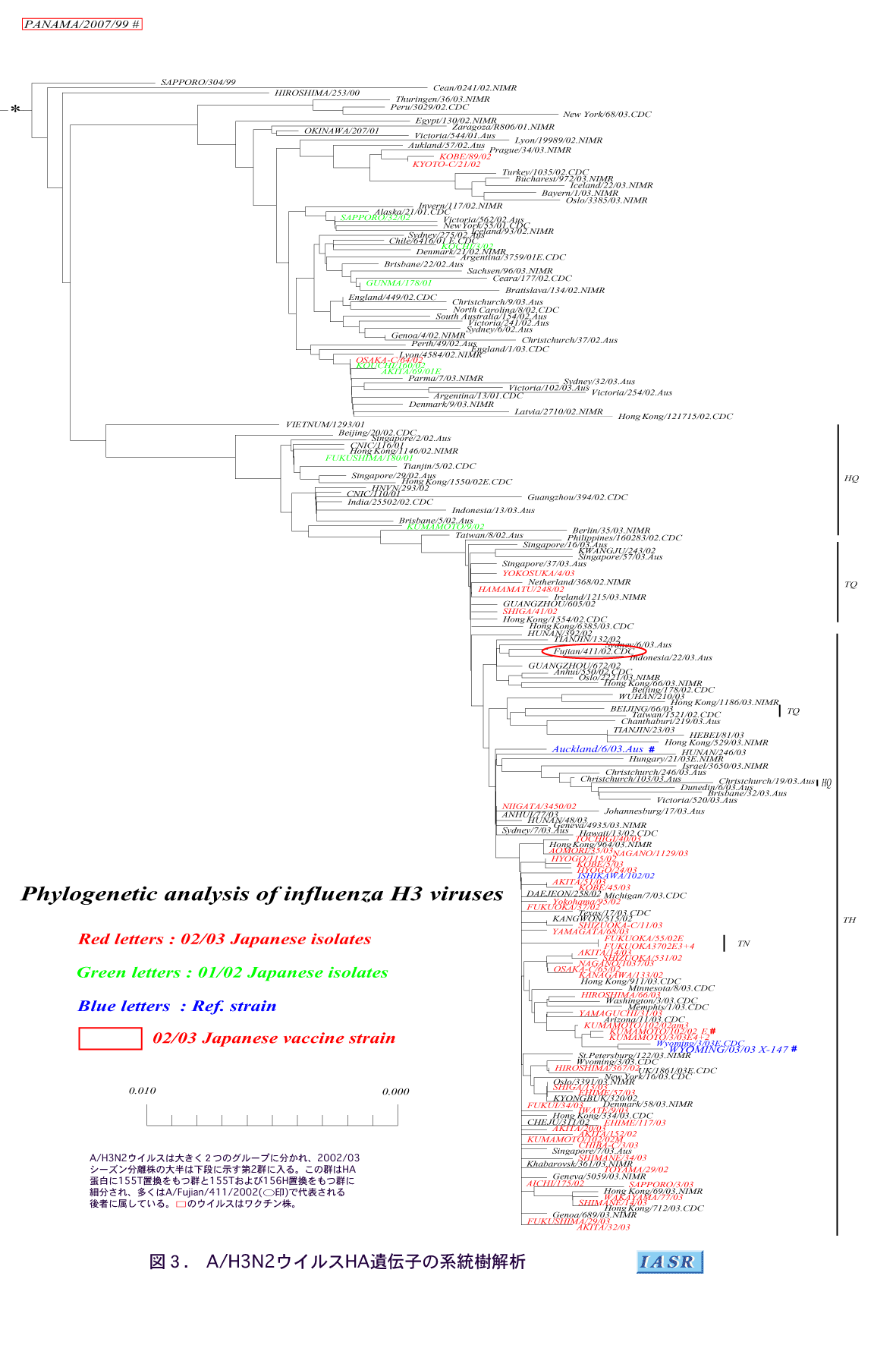
2)A/H3N2ウイルス:A/H3N2ウイルスのHA遺伝子の系統樹は大きく2群に分けられ、 2002/03シーズン分離株の多くは、 前シーズンに東アジア地区で分離された株[A/Hong Kong(香港)/1550/2002、 A/Guangzhou(広州)/394/2002 など]が入る1群から派生していた(図3)。これら分離株は前述したように、 HA蛋白の155のみにアミノ酸置換をもつ群(TQ群)と155および156の2カ所に置換をもつ群(TH群)に細分され、 多くの変異株はA/Fujian/411/2002、 A/Kumamoto/102/2002およびA/Wyoming/3/2003で代表される後者に属していた。
3)B型ウイルス:B型ウイルスは系統樹上からも前述したようにVictoria系統と山形系統に大別される(図4)。Victoria系統のウイルスはB/HongKong/330/2001(丸印)で代表される群とワクチン株B/Shandong/7/97を含むB/Brisbane/32/2002(丸印)で代表される群に分かれるが、 2002/03シーズンの分離株は両方に入り、 これら2群の間では抗原的違いは見られなかった(表3、 図4)。さらに、 ノイラミニダーゼ(NA)遺伝子の塩基配列の決定から、 両群に入る多くの分離株のNA遺伝子は2001/02シーズンのワクチン株B/Johannesburg/5/99を含む山形系統株から由来していることが明らかになっている(図4およびWHO, WER, 78, 57-64, 2003参照)。このことは、 異なる2つの系統間で遺伝子再集合が起こったことを示唆している。
一方、 分離数の少なかった山形系統株は、 B/Johannesburg/5/99、 B/Sichuan(四川)/379/99で代表される群と、 B/Hiroshima/23/2001、 B/Foshan(佛山)/172/2001で代表される2つの群に分かれる(図4)。2002/03シーズンの分離株は後者に入り、 これらはB/Johannesburg/5/99とは抗原的に大きく異なっていた(図2)。
本研究は「厚生労働省感染症発生動向調査に基づくインフルエンザサーベイランス」事業として全国74地研から送付されたウイルス株について、 感染研ウイルス3部第1室(インフルエンザウイルス室)西藤岳彦、 小渕正次、 斉藤利憲、 伊東玲子、 中矢陽子、 板村繁之、 今井正樹、 二宮 愛、 金子睦子、 小田切孝人らにより行われた。また、 本稿に掲載した成績は全解析成績の中から抜粋したものであり、 残りの成績は既にWISH-NETで各地研に還元された。本稿は上記研究事業の遂行にあたり、 地方衛生研究所全国協議会と感染研との合意事項に基づく情報還元である。
国立感染症研究所ウイルス第3部第1室・WHOインフルエンザ協力センター
 | 
| |
| 図3(pdf版) | 図4(pdf版) |
 今月の表紙へ戻る
今月の表紙へ戻る
 IASRのホームページに戻る
IASRのホームページに戻る Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}