2006擭係寧乣2007擭俁寧枛傑偱偵丄晉嶳導偱廤抍敪惗偟偨僂僀儖僗惈堓挵墛偺帠椺偱丄僲儘僂僀儖僗偑専弌偝傟偨傕偺偼28帠椺偁偭偨丅堚揱巕宆偼丄侾椺偑GI/8偱偁偭偨埲奜偼偡傋偰GII/4偱偁偭偨丅偙傟傜偺GII/4偵傛傞帠椺偺側偐偱丄栺俀僇寧娫暢曋拞偵僂僀儖僗堚揱巕偑専弌偝傟丄峔憿抈敀椞堟偺墫婎攝楍偵宱帪揑曄壔偑擣傔傜傟偨帠椺傪宱尡偟偨丅
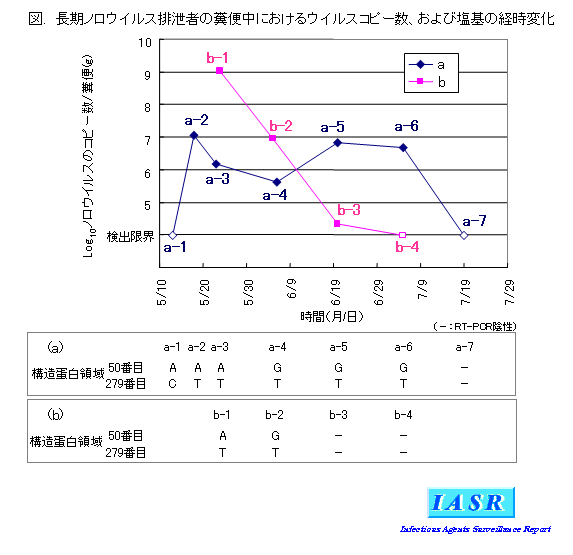
摨帠椺偵偍偄偰丄俀柤偺暢曋偐傜俀僇寧偁傑傝偵傢偨偭偰僂僀儖僗堚揱巕偑専弌偝傟偨丅侾柤(a) 偱偼俈夞専曋傪峴偄丄儕傾儖僞僀儉PCR偵傛傝僂僀儖僗偺僐僺乕悢傪應掕偟偨偲偙傠丄俀曯惈偺宱帪曄壔傪帵偟偨乮恾乯丅傕偆堦椺(b) 偱偼係夞専曋傪峴偄丄摨條偵専摙偟偨偲偙傠丄僂僀儖僗偺僐僺乕悢偼姶愼弶婜偵旕忢偵懡偔丄偦偺屻媫寖偵尭彮偡傞宱夁傪帵偟偨乮恾乯丅RT-PCR丄偍傛傃僟僀儗僋僩僔乕僋僄儞僗偵傛傝僂僀儖僗堚揱巕偺墫婎攝楍傪夝愅偟偨寢壥丄峔憿抈敀椞堟偺奐巒僐僪儞偐傜悢偊偰50斣栚偺墫婎偑丄(a) 偱偼丄俁夞栚偲係夞栚偺専懱偺娫偱俙偐傜俧偵曄壔偟偰偄傞偙偲偑敾柧偟偨乮恾乯丅(b) 偱傕丄侾夞栚偲俀夞栚偺専懱偺娫偱丄摨條偵俙偐傜俧偵曄壔偟偰偄偨乮恾乯丅宱夁拞偵僂僀儖僗堚揱巕偑曄堎偟偨壜擻惈偑峫偊傜傟偨偨傔丄PCR嶻暔傪僋儘乕僯儞僌偟偰丄屄乆偺僋儘乕儞偺墫婎攝楍傪専摙偟偨丅偦偺寢壥丄a-1乣a-3偱偼偡傋偰偺僋儘乕儞偑俙攝楍傪帩偭偰偄偨偺偵懳偟偰丄a-4偱偼24僋儘乕儞拞俙偑俋丄俧偑15丄a-5偱偼俙偑侾丄俧偑24丄a-6偱偼偡傋偰俧偱偁偭偨丅傑偨丄b-1偱偼丄28僋儘乕儞偡傋偰偑俙偩偭偨偺偵懳偟偰丄b-2偱偼丄29僋儘乕儞拞俙偑侾丄俧偑28僋儘乕儞偩偭偨丅偙偺傛偆偵丄俙傪帩偮僂僀儖僗偐傜俧傪帩偮僂僀儖僗傊偲妱崌偑曄壔偟偰偄傞偙偲偑敾柧偟偨丅偙偺墫婎偺曄壔偵傛傝丄峔憿抈敀偺17斣栚偺傾僗僷儔僊儞乮CAA乯偑僙儕儞乮CAG乯偵曄壔偡傞偙偲偑傢偐偭偨丅忋婰偺曄壔偵壛偊偰(a) 偱偼丄峔憿抈敀椞堟偺奐巒僐僪儞偐傜悢偊偰279斣栚偺墫婎傕丄侾夞栚偲俀夞栚偺専懱偺娫偱俠偐傜俿偵曄壔偟偰偄傞偙偲偑敾柧偟偨乮恾乯丅摨條偵PCR嶻暔傪僋儘乕僯儞僌偟偰妱崌傪媮傔偨偲偙傠丄a-1偱偼偡傋偰偑俠丄a-2偱偼偡傋偰偑俿偲丄偙偺娫偵摨晹埵偺墫婎偑堦曄偟偰偄傞偙偲偑傢偐偭偨丅(b) 偱偼偡傋偰俿偱曄壔偼擣傔傜傟側偐偭偨丅偙偺俠偐傜俿傊偺曄壔偼丄傾儈僲巁偺曄壔傪敽傢側偄偙偲偑傢偐偭偨丅屄乆偺僋儘乕儞偵偼丄忋婰俀僇強埲奜偺晹埵偱偺墫婎偺曄壔傕嶶尒偝傟偨偑丄偄偢傟傕屌掕壔偡傞偙偲偼側偐偭偨丅
(a) 丄(b) 俀椺偱丄摨偠曄堎偑摨帪偵婲偒傞壜擻惈偼嬌傔偰掅偄偲峫偊傜傟傞偨傔丄堦曽偱惗偠偨墫婎曄壔傪傕偮僂僀儖僗偑懠曽偵姶愼偟偨壜擻惈偑崅偄偲峫偊傜傟傞丅(a) 偺僂僀儖僗検偺宱帪曄壔偑俀曯惈傪帵偟丄墫婎偺妱崌曄壔傕偙傟傜僺乕僋偺忋徃慜偵婲偒偰偄傞偙偲偐傜丄偦傟偧傟偺捈慜偵(b) 偐傜(a) 偵姶愼偟偨壜擻惈偑峫偊傜傟傞丅
宱夁拞偵堚揱巕曄壔偑擣傔傜傟偨尨場偲偟偰丄挵娗偱僂僀儖僗偑憹怋偡傞娫偵曄堎偑婲偒偨丄堎側傞僂僀儖僗偑嵞姶愼偟偨丄傕偲傕偲彮検崿嵼偟偰偄偨僂僀儖僗偑桪惃偵側偭偨偙偲側偳偑峫偊傜傟傞丅(b) 偺姶愼弶婜偵偼僂僀儖僗偺僐僺乕悢偑旕忢偵懡偐偭偨偙偲偐傜丄僂僀儖僗偺憹怋偑惙傫偱丄偦偺娫偵僂僀儖僗偑曄堎偟偨壜擻惈傕峫偊傜傟傞丅
俙偐傜俧偺曄壔偵傛傝傾僗僷儔僊儞偑僙儕儞偵曄壔偡傞偙偲偐傜丄峔憿抈敀偺峈尨惈偑曄壔偟偨壜擻惈傕峫偊傜傟傞偑丄摨晹埵偵偼柧妋側峈尨寛掕晹埵偼曬崘偝傟偰偄側偄丅傑偨丄俠偐傜俿傊偺曄壔偼傾儈僲巁曄壔傪敽傢側偄丅偟偨偑偭偰丄峈懱偺塭嬁偵傛傝偙傟傜偺曄壔偑(a) 偱桪惃偵側傝屌掕壔偟偨偲偼峫偊偵偔偄丅僂僀儖僗堚揱巕偺懠偺椞堟偵惗偠偨曄堎偵傛傞塭嬁偺壜擻惈傕峫偊傜傟丄崱屻偺専摙偑昁梫偱偁傞丅
杮帠椺偵傛傝丄挿婜僲儘僂僀儖僗攔煏幰拞偵偍偗傞僂僀儖僗曄堎偺壜擻惈偑帵嵈偝傟偨丅挿婜攔煏椺偱偼丄堚揱巕偑宱夁拞偵曄壔偡傞壜擻惈傪峫偊偰丄崱屻専摙傪廳偹偰偄偔昁梫偑偁傞偩傠偆丅
晉嶳導塹惗尋媶強僂僀儖僗晹
彫尨恀媩丂挿扟愳悷戙丂拞懞堦嵠丂娾堜夒宐丂杧尦塰帉丂憅揷丂婤丂戧郪崉懃
崅壀岤惗僙儞僞乕丂陰摗彯恗
怴愳岤惗僙儞僞乕丂戝峕丂峗
 崱寧偺昞巻傊栠傞
崱寧偺昞巻傊栠傞
 IASR偺儂乕儉儁乕僕偵栠傞
IASR偺儂乕儉儁乕僕偵栠傞 Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}