破傷風毒素の構造
破傷風毒素は破傷風菌(Clostridium tetani )の増殖とともに菌体内に産生され、対数増殖期の後期から培養上清中に放出される分子量約15万の単純蛋白質である。菌体内で1315のアミノ酸からなる一本鎖のポリペプチドとして合成された毒素は、菌体外に放出される際に457番目のAlaから461番目のAspまでの間でトリプシン様の蛋白質分解酵素によって限定分解を受け、N端側の分子量約5万の軽鎖(Light chain)とC端側の分子量約10万の重鎖(Heavy chain)の二本鎖がジスルフィド結合によって架橋された4次構造を形成する。軽鎖と重鎖との結合は強固で毒素をDTTなどの還元剤で処理するだけでは解離せず、2M尿素などの変性剤で処理することで重鎖と軽鎖をそれぞれ分離することができる1)。また破傷風毒素を穏やかな条件下でパパイン処理することで、C端側5万のペプチドフラグメントとN端側10万のフラグメントに分けることができることから、重鎖のC端側5万をFragment C(Frg C)、N端側5万をFragment B(Frg B)、さらに軽鎖をFragment A(Frg A)と呼称することが第8回国際破傷風会議(1987)で採択された。一方、破傷風菌と同属のボツリヌス菌が産生するボツリヌス毒素と破傷風毒素の構造的類似点が明らかになるに従って、ボツリヌス毒素で提唱されている構造名称をそのまま破傷風毒素にも適応する研究グループも現れている。図1に破傷風毒素とボツリヌス毒素の模式的構造と部分名称の対比を示す。それぞれの神経毒素の3Dモデルはhttp://www.ncbi.nlm.nih.gov/sites/entrezからダウンロードできる。遺伝子レベルでの比較では破傷風毒素と各血清型のボツリヌス毒素との相同性は低い。また、ペプチドレベルでも破傷風毒素のFrg BはB型ボツリヌス毒素のHN に比べて疎水性が極めて高く、尿素を透析で除くと容易に不溶化沈殿するという違いがある。3Dモデルで示された全体像の類似性に注目するか、部分的な差異に注目するかについては意見の分かれるところである。
破傷風毒素の分子内機能分担
Fragment A:破傷風毒素の軽鎖(Frg A)のほぼ中央(233-237 AA)に亜鉛結合構造HEXXHがあることが破傷風毒素遺伝子の塩基配列の決定によって明らかとなり2)、1992年Schiavo G 3)らはシナプス小胞の膜蛋白質Synaptobrevinが破傷風毒素の軽鎖によって限定分解を受けることを報告した。これによって破傷風毒素のFrg Aがメタロプロテアーゼであり、基質であるvSNAREが限定分解を受けることでシナプス小胞とシナプス前膜とのドッキングが阻害される。これが破傷風毒素によるシナプス前抑制の分子機構であることが明らかになった。
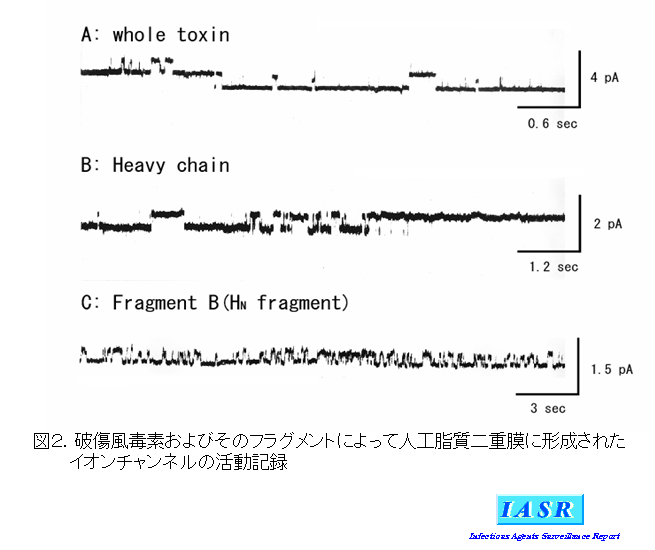
Fragment B:破傷風毒素の重鎖N端側のFrg Bは疎水性が極めて高く、容易に不溶化沈殿するためにその機能の解析が進んでいない。1989年Matsuda M 4)らはパパイン処理した破傷風毒素から精製したFrg Bが人工脂質二重膜にイオン透過性チャンネルを形成する機能があることを報告した。図2に人工脂質二重膜に形成されたイオンチャンネルの記録を示す。口径が約1μmのガラスピペットの先端にリン脂質による人工二重膜を作製し、破傷風毒素側のpHを4.5に下げると、図2Aに示した電流変化が記録される。電流の変化はランダムに生じるがレベルは一定であることから、イオンチャンネルの活動であることが分かる。このチャンネル形成活性は重鎖(図2B)およびFragment B単独(図2C)でも見られることから、Frg B部分に担われていると推測された。チャンネル形成活性が毒素作用の発現機構に果たす役割については不明な点が多いが、ジフテリア毒素のフラグメントBもチャンネル形成活性を有していることから、酵素活性を有しているFrg Aがエンドゾームから細胞質に移行する過程に関与していると考えられている。
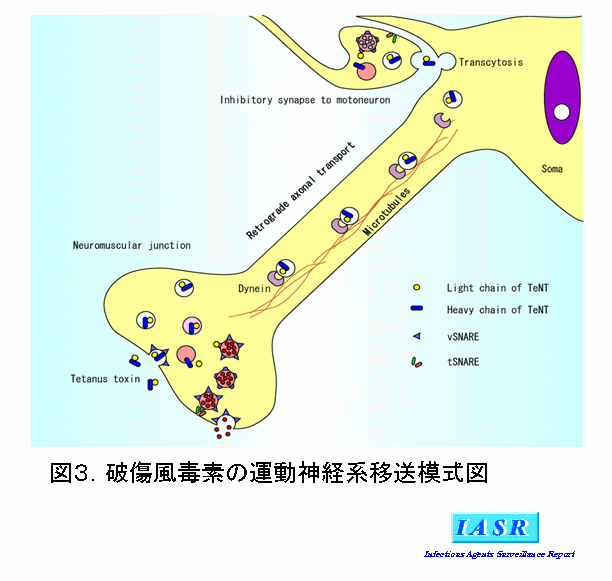
Fragment C:重鎖C端側のFrg Cについては受容体結合部位であろうと推測され、Frg Cを用いて破傷風毒素の受容体を同定する試みが続けられている。Frg CがGT1bやGD1bなどのPolysialogangliosideと会合することは間違いないが、破傷風毒素の極めて高い神経向性はガングリオシドが毒素受容体では説明できず、多くの研究者は蛋白質の関与を想定している。Herreros J 5)らはThy-1が毒素受容体の蛋白質成分である可能性を指摘したが、後にThy-1ノックアウトマウスの破傷風感受性に変化がないことが示された。現在もなお破傷風毒素の受容体は不明のままであるが、Lipid raftに存在するGPI-anchored蛋白質に注目して検索を進めているグループもある。破傷風毒素の運動神経終末部への取り込みについても、シナプス小胞のリサイクリング・lipid raft・clathrin-coated pitなどとの関係について諸説があり、不明のままである。神経終末部での取り込みの後、破傷風毒素が中枢の神経細胞体へ逆行性に運ばれ、さらにシナプスを越えて高次神経細胞のシナプス前部に到達する過程(Transcytosis)にFrg Cが関与していることが、Rind HBら 6)およびBohnert Sら 7)によって示された。標識されたFrg Cは取り込まれた後も中性を保ったコンパートメントで細胞体へと逆行性に運搬され、運動神経の樹状突起に集積される。BDNFやGDNFなどの神経栄養因子と比較すると、運搬速度や樹状突起への集積速度は同じ(ca. 1μm/sec)であるが、シナプスを越えて次のシナプス前部への移行はFrg Cの方がほぼ倍の速度で行われることが明らかにされた。
破傷風毒素の蛋白質としての構造および機能分担について現在までに解明されたことについて述べた。破傷風毒素の研究は、1890年にKitasato Sによる発見以来百年以上経過した。しかし我々が現在知っていることはといえば、蛋白質としての毒素と酵素活性だけである。破傷風毒素の神経毒素としての特異性を決定する事柄、1)毒素受容体、2)毒素の取り込み、3)逆行性軸索輸送へのソーティング、4)シナプスを越えてのtranscytosis、5)Frg Aの細胞質への移送(図3)、は現在もなお不明のまま残されている。
文 献
1) Matsuda M and Yoneda M, Infect Immun 12: 1147-1153, 1975
2) Fairweather NF and Lyness VA, Nucleic Acids Res 14: 7809-7812, 1986
3) Schiavo G, et al ., Nature 359: 832-835, 1992
4) Matsuda M, et al ., Infect Immun 57: 3588-3593, 1989
5) Herreros J, et al ., Mol Biol Cell 12: 2947-2960, 2001
6) Rind HB, et al ., J Neurosci 19: 539-549, 2005
7) Bohnert S and Schiavo G, J Bio Chem 280: 42336-42344, 2005
大阪大学医学系研究科・感染防御学 杉本 央
 今月の表紙へ戻る
今月の表紙へ戻る
 IASRのホームページに戻る
IASRのホームページに戻る Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}
{kind=link}
{kind=link}