�A�V�l�g�o�N�^�[���ۂ́A�Δ^�ۂȂǂƓ��l�̃O�����A�����ۂɑ����A�^�����͂Ȃ��B�ʏ�́A�u�h�E�������C�I�����ŕ����i���y�j���Ȃ����A�A�V�l�g�o�N�^�[���ۂ̒��ŁA��q����Acinetobacter baumannii �́A���̎�ƈقȂ�A�u�h�E����L�_�f���Ŏ_���I�ɕ������邱�Ƃ��ł���B�A�V�l�g�o�N�^�[���ۂ́A�y��A�͐쐅�Ȃǎ��R�������炵���Ε����������ۂł���A�����ɔ�r�I���������������A�܂������̕��ւȂǂ�������������B�A�V�l�g�o�N�^�[���ۂ́A�ۑ̊O�ɑ��݂���DNA�f�Ђ���荞��ŁA���Ȃ̐��F��DNA�Ȃǂɑg�ݍ��ދ@�\�������߁A�O�����A�����ۂ̒��ł́A���R�`���]�����N�����₷���B
�����Ǝ햼
1950�N��܂ł́AMoraxella lwoffii �ƌĂ�Ă����ێ�́A1960�N�����AAcinetobacter lwoffii �Ƒ������ύX���ꂽ�B�܂��AAcinetobacter calcoaceticus ��1970�N����V������Ƃ��ĔF�m�����悤�ɂȂ�A���݁A�Տ�����Ŗ��ƂȂ��Ă���A. baumannii �́A1980�N����l�ɕa������������Ƃ��ĐV���ɔF�m�����悤�ɂȂ����B���݁A�A�V�l�g�o�N�^�[���ɂ́A���Ȃ��Ƃ�17�̎햼��15��genomic species���m�F����Ă���B
���ܑϐ����̑ϐ��@�\
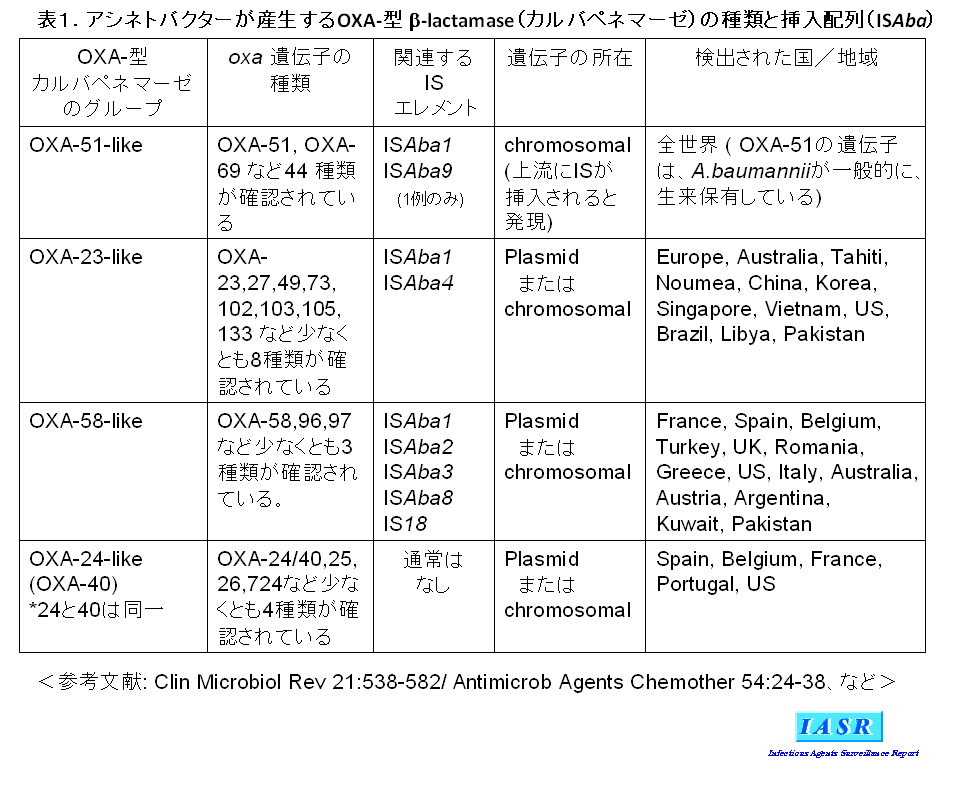
�L��Z�t�@���X�|�����ϐ��ɂ͐��F�̏��AmpC�^�Z�t�@���X�|���i�[�[�iADC-1�Ȃǁj�̎Y�����֗^���邪�A�J���o�y�l���ϐ��ɂ́AA. baumannii �����������Ă���OXA�^�J���o�y�l�}�[�[OXA-51-like�̈�`�q�̏㗬�Ƀv�����[�^�[������L����}���z��iISAba1 �Ȃǁj���}�����ꂽ��A�O������OXA-23-like��OXA-58-like�Ȃǂ̃J���o�y�l�}�[�[�̈�`�q�̊l�����֗^���Ă���i�\�P�j�B
�t���I���L�m�����ϐ��ɂ́A�Δ^�ۂȂǂƓ��l�ɁA���F�̏�ɑ��݂���DNA �W���C���[�X��g�|�C�\�����[�[IV�Ȃǂ�DNA�����y�f�̃L�m�����ϐ�����̈�iQRDR�j�̃A�~�m�_�c��̒u���������N������`�q�ψق�R�ۖ�r�o�@�\�iAdeABC�j�Ȃǂ��֗^����B
�A�~�m�z���̑ϐ��ɂ��Ă��A�Δ^�ۂȂǂƓ��l�ɃA�~�m�z���̂̃����_���y�f�iAPH�j��A�Z�`�����y�f�iAAC�j�A�A�f�j�������y�f�iAAD�j�Ȃǂ̎Y�����֗^����B���ɁA�A�~�m�z���̂̕W�I���q�ł���16S rRNA�����`��������y�f�iArmA�j���Y�����A�L�͈͂̃A�~�m�z���̂ɒ����x�ϐ����l����������������č��Ȃǂő���������A�ŋ߁A��t���ł�ArmA�Y�������m�F����Ă��邱�Ƃ���A����̍L���肪�x������Ă���B
���������nj������ۑ�@�r��X�e
 �����̕\���֖߂�
�����̕\���֖߂�
 IASR�̃z�[���y�[�W�ɖ߂�
IASR�̃z�[���y�[�W�ɖ߂� Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}