肺炎マイコプラズマ(Mycoplasma pneumoniae )の培養検査は手技が煩雑な上に日数を要することから、国内でこれを実施している機関は極めて少ない。神奈川県衛生研究所では、1976年から培養検査によるM. pneumoniae 感染症の調査をしており、その中で培養検査法やPCR法の検討をし、さらに分離株を使用した薬剤感受性試験、P1蛋白遺伝子型別等を実施してきた。今回は、当所で実施している培養検査法、PCR法およびP1蛋白遺伝子型別法、ならびにそれらの成績について報告する。
1.培養検査
検査法:M. pneumoniae 感染を疑われた患者の咽頭スワブを下記の液体培地2ml中で丹念に絞り出し、その絞り液を培養用検体とした。培養検査には、BBLTM Mycoplasma broth base (B.D.) 、ウマ血清(56℃ 30分加熱、Gibco)20%、酵母エキス(自家製)10%、ブドウ糖1%、ペニシリン-G 103 U/ml、酢酸タリウム0.025%の組成から成るものを液体培地として使用した。これらにBactoTMagar(B.D.)1.5%を加えて寒天培地とし、平板培地および二層培地の作製に使用した。二層培地はフェノールレッド0.002%およびメチレンブルー 0.001%を添加した寒天培地と液体培地を使用して作製した選択分離培地で、前者1mlに後者2mlを重層したものである。以上の培地の他に、市販生培地であるマイコプラズマPPLO寒天培地(日研生医研)を分離平板培地として併用した。これらの平板培地および二層培地に咽頭スワブ絞り液を接種後、37℃で好気培養した。M. pneumoniae の同定は、PCR(下述)あるいは抗血清による発育阻止試験で行った。
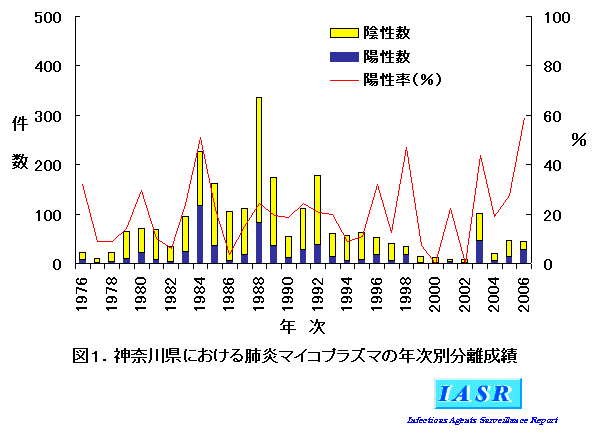
検査成績:神奈川県において、1976~2006年までに検査した咽頭スワブ2,414件中、M. pneumoniae 培養陽性は585件(陽性率24%)であった。図1に年次別成績を示した。1988年までは培養陽性数と陽性率がともにピークを示す現象が4年ごとに見られたが、以後はその周期性は崩れている。これはわが国におけるM. pneumoniae 感染症の発生動向とほぼ一致している。昨年(2006年)は全国的にM. pneumoniae 感染症が多発したことが報告されており(本号特集参照)、本県においても培養陽性率は59%と高い値が示された。
なお、従来、M. pneumoniae 培養検査用培地の基礎培地として使用していたBacto PPLO broth(Difco)が入手困難となったため、現在はBBLTM Mycoplasma broth base (B.D.)を使用しているが、特に問題は起きていない。また、市販生培地であるマイコプラズマPPLO寒天培地(日研生医研)は、分離培地として十分に使用可能と思われる。しかし、M. pneumoniae のコロニー形態が他の寒天培地とはやや異なるので注意が必要である。
2.M. pneumoniae のPCR法による検出
PCR法:M. pneumoniae の16S rRNAやP1蛋白遺伝子等の特異領域を標的とするプライマーがいくつか報告されているが、当所ではIevenら(J Infect Dis, 1996)が記載した16S rRNA遺伝子を標的としたプライマーを使用している。PCR 用検査材料には上述の培養検査に使用する咽頭スワブ絞り液を使用した。この1mlを遠心(15,000rpm 20分)し、沈渣にSTE 緩衝液0.1mlを加えた後、 100℃10分加熱してテンプレートDNAを作製した。増幅反応は、94℃2分加熱後、94℃1分、55℃1.5分、72℃1分を40サイクルで行い、増幅産物を2%アガロースゲルにて電気泳動した。
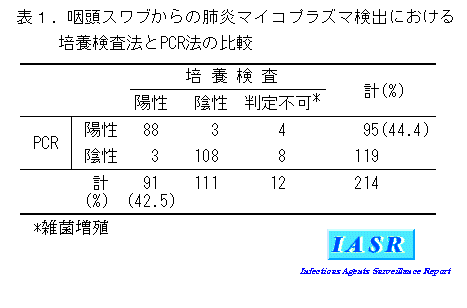
PCRと培養検査法の比較:神奈川県内において、2003~2006年に採取された214件の咽頭スワブにつき、PCRと培養検査の両者を実施した結果を表1に示す。両者の陽性率に差はほとんど見られなかった。一方、雑菌増殖により培養検査では判定できなかった12件(5.6%)のすべてがPCR法で判定され、しかも4件が陽性であった。また、これらの12件を除き、培養検査法を確定診断法として算出したPCRとの陽性一致率は96.7%、陰性一致率は97.3%となり、両者の成績は良く一致した。これらの結果は、M. pneumoniae 感染症の診断におけるPCR法の有用性を示すものである。今後、現在報告されているいくつかのプライマーについて特異性および感度を比較した上で、より有用なPCR法を確立していく必要があると考えられる。
3.P1蛋白遺伝子型別法
型別法:M. pneumoniae のP1蛋白遺伝子の前半部と後半部の可変領域をPCRで増幅し、増幅産物を制限酵素Hae IIIで切断後、アガロース電気泳動して切断パターンを観察する(Sasakiら、J Clin Microbiol, 1996)。M. pneumoniae の液体培養1mlの遠心沈渣(15,000rpm 10分)に0.1mlの精製水を加えて100℃10分加熱し、テンプレートDNAを抽出した。PCRは、ADH1、ADH2およびADH3、ADH4の2組のプライマー(Sasakiら、J Clin Microbiol, 1996)を使用し、94℃1分加熱、94℃1分、55℃1分および72℃ 2.5分を30サイクルで行った。PCR増幅産物40μlに3M酢酸ナトリウム(<pH 5.2)を4μl、純エタノール80μlを加えて混合し、遠心後(10,000rpm、10分)、-20℃に冷却しておいた70%エタノールで沈殿物を洗浄した。その後、デシケーターで乾燥し、精製水20μlを加えた。これに制限酵素Hae III、緩衝液および精製水を加えて50μlとし、37℃90分反応させた。制限酵素処理後、3%アガロースにより電気泳動し、切断パターンを観察した。
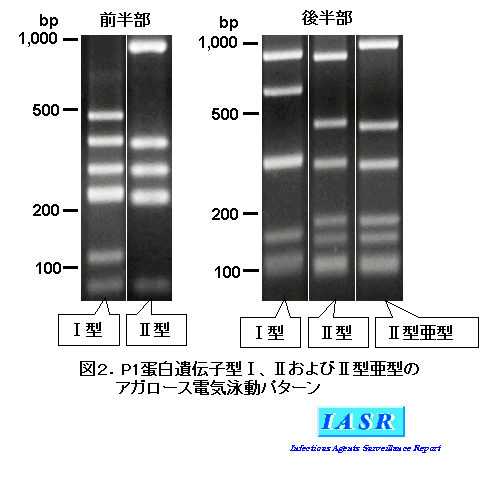
電気泳動パターン:現在、P1蛋白遺伝子型としてIおよびII型が知られており、II型には亜型が存在する(Kenriら、Infect Immun, 1999)。図2に、それらのアガロース電気泳動パターンを示した。P1蛋白遺伝子前半部および後半部ともにI型パターンを示す菌をI型、同様にII型パターンを示す菌をII型とし、前半部はII型パターンで、後半部がI型でもII型パターンでもない菌をII型亜型としている。わが国では、IおよびII型菌が一定期間をもって入れ替わる現象が見られる。最近の数年間はI型菌が優勢であり、II型菌のほとんどがII型亜型であることが特徴である(本号特集参照)。
神奈川県衛生研究所微生物部 岡崎則男 大屋日登美
国立感染症研究所細菌第二部 佐々木次雄 見理 剛
 今月の表紙へ戻る
今月の表紙へ戻る
 IASRのホームページに戻る
IASRのホームページに戻る Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}
{kind=link}
{kind=link}