2008/09インフルエンザシーズンは2008年第45〜46週(11/3〜11/16)のA/H1N1、A/H3N2亜型およびB型株の混合流行で幕を開け、2009年第4週(1/19〜1/25)にA/H1N1亜型を主流としたピークが見られた。A型の流行はその後急速に減少し、第13週(3/23〜3/29)にはほぼ終息した。一方、B型の流行はA型より低レベルで始まり、第10〜11週(3/2 〜3/15)をピークとして第21週(5/18〜5/24)には終息した。
一方、3月中旬にメキシコで発生したブタ由来のA/H1N1新型インフルエンザはその後も流行を広げ、4月中旬には米国南カリフォルニアへ波及した。その後、瞬く間に世界中に広がり、6月11日にはWHO から『パンデミック宣言』に相当するパンデミック警戒レベルのフェーズ6が出された。本邦では、遡り疫学調査によって、5月3日神戸市での初発例が確認されているが、最初のウイルス分離は、5月8日に成田空港検疫所で捕捉されたカナダから帰国した大阪の高校生グループの患者からのもので、分離株はA/Narita(成田)/1/2009(H1N1)pdmと命名された。それ以降、関西地区で相次いで新型インフルエンザウイルスが分離され、6月以降は全国各地でもウイルスが検出されるようになった。第33週(8/10〜8/16)には定点医療機関での週当たりのインフルエンザ様患者報告数が1.0を超え、異例ではあるが夏場のインフルエンザ流行となった。第20週(5/11〜5/17)以降は少数のA/H3N2亜型およびB型ウイルスとの混合流行であったが、第28週(7/6〜7/12)以後では、分離株のほぼ98%は新型インフルエンザA(H1N1)pdmで占められるようになった。
2008/09シーズン最終の第35週までの総分離数9,962株における分離比は、季節性A/H1N1が33%、A/H3N2が18%、B型が19%、新型A(H1N1)pdmが29%であった。
2.ウイルス抗原性および遺伝子系統樹解析
2008/09シーズンに全国の地方衛生研究所(地研)で分離されたウイルス株は、各地研において、国立感染症研究所(感染研)からシーズン前に配布された抗原性解析用抗体キット[A/Brisbane/59/2007(H1N1)、A/Uruguay/716/2007(H3N2)、B/Malaysia/2506/2004(Victoria系統)、B/Brisbane/3/2007(山形系統)]を用いた赤血球凝集抑制(HI)試験によって、型・亜型同定および初期抗原性解析が行われた。感染研では、これらの成績を感染症サーベイランスシステム(NESID)経由で収集し、HI価の違いの比率が反映されるように選択した分離株(分離総数の約5〜10%に相当)および非流行期の分離株や大きな抗原性変化を示す分離株の分与を地研から受け、それらについてフェレット参照抗血清を用いて詳細な抗原性解析を実施した。
2-1)A/H1N1ウイルス
抗原性解析:国内では3,302株のAH1亜型ウイルスが全国の地研で分離された。感染研ではこれらの国内および海外(中国、台湾、韓国、ラオス、モンゴル、ミャンマー)から収集した総数337株について、5種類のフェレット参照抗血清を用いて抗原性解析を行った。
シーズン前半(2008年9月〜2009年2月)および後半(2009年3月〜8月)の分離株のそれぞれ94%、85%は、参照株でワクチン株にも選定されたA/Brisbane/59/2007と抗原性が類似していた。一方、HI試験で抗A/Brisbane/59/2007血清から8倍以上減少した変異株も少数ながら検出されたが、これらが増える傾向はシーズンを通して見られなかった。
諸外国においても新型インフルエンザA(H1N1)pdmウイルスの流行が始まるまでは、季節性A/H1N1ウイルス(ソ連型)の流行が主流であり、その抗原性はわが国と同様にA/Brisbane/59/2007類似株が大半を占めていた。
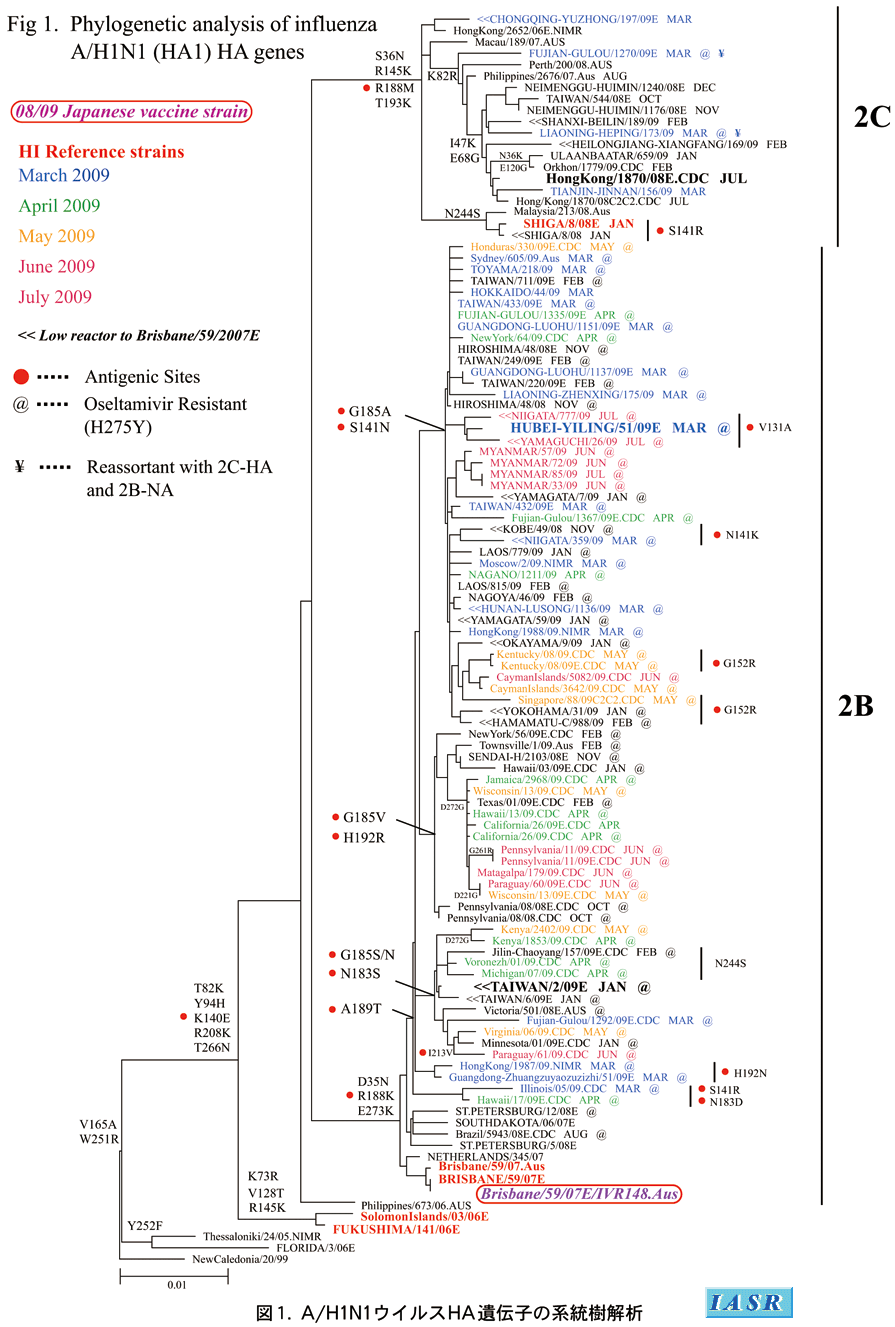
遺伝子系統樹解析:HA遺伝子の系統樹解析では、国内分離株のすべては、D35N、R188K、E273Kのアミノ酸置換をもち、その代表株であるA/Brisbane/59/2007が入る一群(クレード2B)に分類された(図1)。このクレード2Bのウイルスは、ノイラミニダーゼ(NA)蛋白にオセルタミビル(タミフルR)耐性のマーカーH275Y をもち、感受性試験においてもオセルタミビル耐性であることが確認された(3.抗インフルエンザ薬耐性株の項参照)。
一方、中国での分離株の多くは、S36N、R145K 、R188M、A189T、T193Kのアミノ酸置換をもち、その代表株のA/Hong Kong(香港)/1870/2008を含む群(クレード2C)に分類された(図1)。これらは、アマンタジンには耐性であるが、オセルタミビルおよびザナミビルには感受性であった。しかし、シーズン後半からは、中国においてもクレード2Cに入る株は減少し、オセルタミビル耐性のクレード2B株が急激に増加する傾向が見られ、今冬は中国でもオセルタミビル耐性株が主流になることが予想された。
2-2)A/H3N2ウイルス
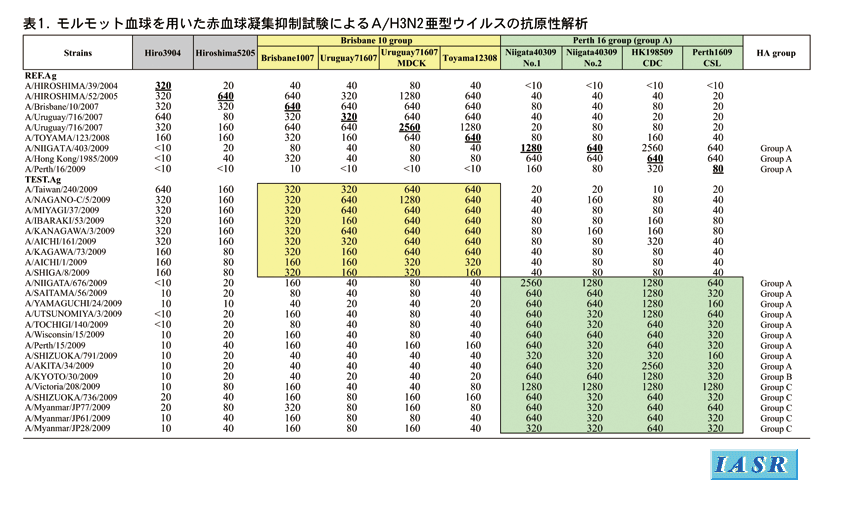
抗原性解析:国内では1,839株のAH3亜型ウイルスが分離された。今シーズンの本亜型による流行は、第3〜6週(1/12〜2/8)の季節性インフルエンザ流行時期と、新型A(H1N1)pdmウイルスとの同時流行期の第20〜23週(5/11〜6/7)にピークをもつ二峰性であった。感染研では国内外から収集した 203株について、8〜10種類のフェレット参照抗血清を用いて抗原性解析を行った(表1)。
シーズン前半では、解析した分離株の72%がワクチン株A/Brisbane/10/2007またはA/Uruguay/716/2007類似株で占められていた。一方、シーズン後半になると、ワクチン類似株は急速に減少し、一方では、HI試験でA/Brisbane/10/2007に対する反応性が8倍以上減少した変異株が出現し、これが分離株の75%を占めるようになった。これら変異株群はA/Perth/16/2009、A/Hong Kong (香港)/1985/2009およびA/Niigata(新潟)/403/2009フェレット抗血清とよく反応し、抗原的にA/Brisbane/10/2007類似株群とは明らかに区別された(表1)。すなわち、本亜型の最初のピークはA/Brisbane/10/2007類似株で、第2のピークはA/Perth/16/2009類似株で起こった流行であった。
諸外国でもシーズン後半にはA/Perth/16/2009類似の変異株が急増し、南北アメリカ分離株の41%、オーストラリアを中心とした南半球分離株の74%およびヨーロッパ地域分離株の31%は変異株であり、世界的にA/Brisbane/10/2007類似株からA/Perth/16/2009類似株へと流行株の主流が移行していく傾向がみられた。
ここ1〜2シーズンのA/H3N2分離株においては、赤血球凝集素(HA)蛋白上のレセプター結合部位の変化が生じて七面鳥赤血球を凝集しないものが急増している。ヨーロッパ諸国の分離株は特に顕著で、多くはモルモット赤血球のみを凝集した。アメリカおよび南半球からも同様の報告がある。国内分離株も現時点では少数ではあるがモルモット赤血球のみを凝集し、七面鳥赤血球ではHI試験ができない傾向が見られている。従って、感染研では、2009/10シーズンのA/H3N2分離株のHI試験は、1%モルモット赤血球を用いて実施することとした。
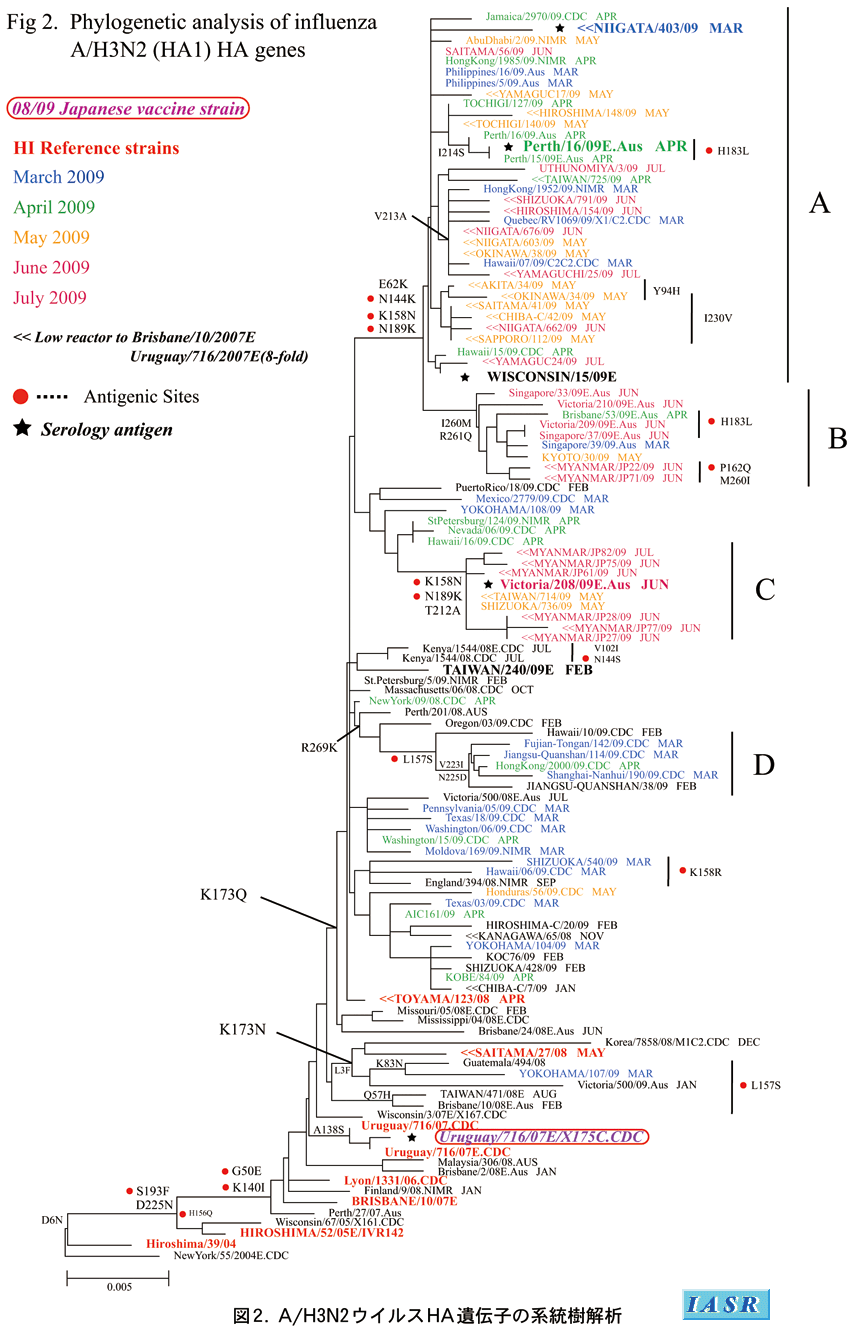
遺伝子系統樹解析:HA遺伝子系統樹ではK173Nのアミノ酸置換を持つクレード1と、K173Qのアミノ酸置換を持つクレード2に分類された(図2)。シーズン全体を通じて国内外の分離株はすべてクレード2に属しており、これらはさらにA/Niigata(新潟)/403/2009やA/Perth/16/2009で代表されるE62K、N144K、K158N、N189K置換を持つグループA、A/Kyoto(京都)/30/2009やA/Singapore/37/2009が入りI260M、R261Q置換を持つグループBおよびA/Victoria/208/2009およびミャンマー株が入りK158N、N189K、T212A置換を持つグループCに大別された(図2)。また、L157S置換を持つグループDも識別されたが、5月以降はこのグループのウイルスは分離されなかった。これらA、B、Cグループに属する分離株のすべては抗原的には、ワクチン株A/Brisbane/10/2007とは大きく異なっており、いずれもA/Perth/16/2009類似株であったが、各遺伝的グループ間では抗原性に差はなかった。
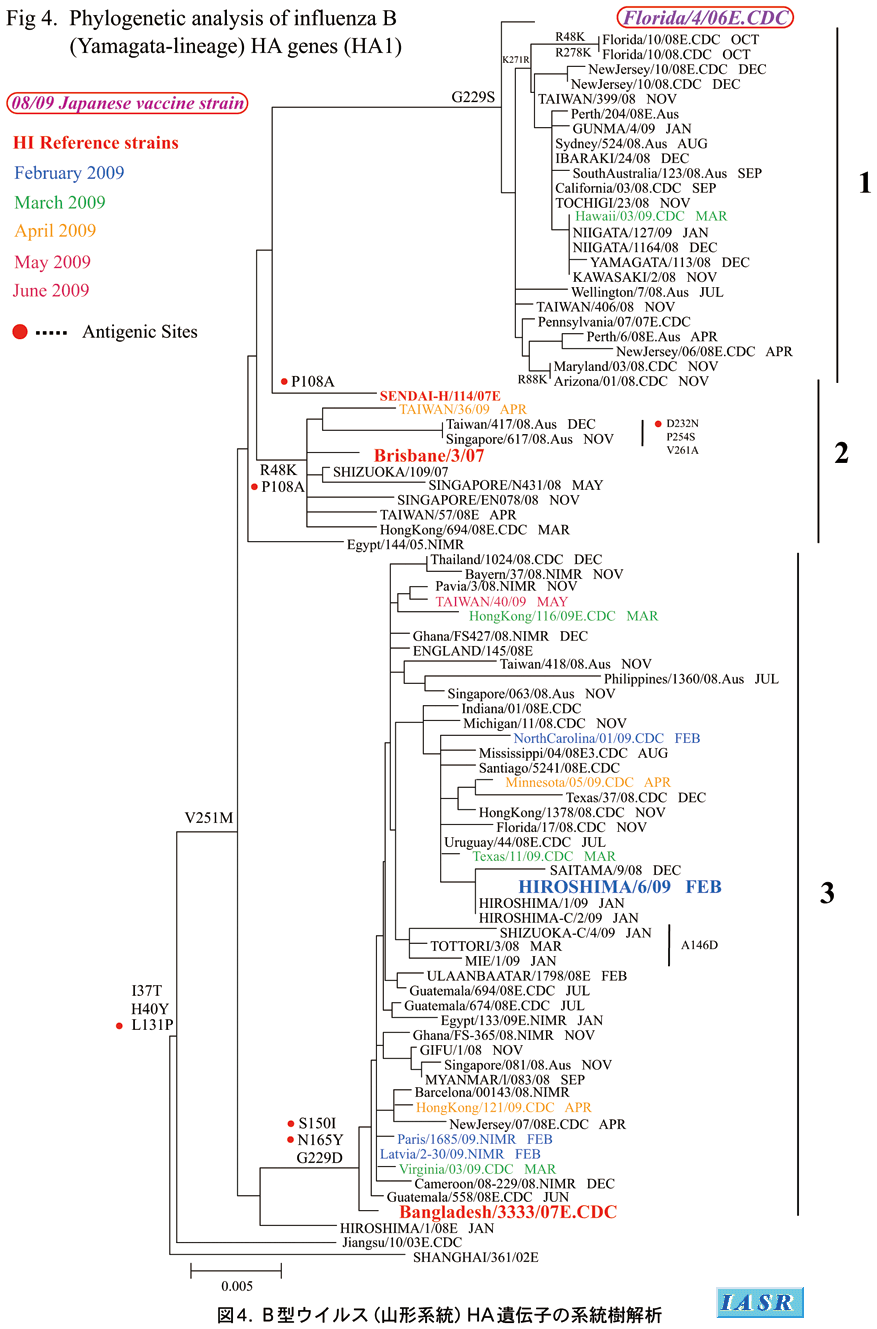
2-3)B型ウイルス
B型インフルエンザウイルスには、B/Yamagata(山形)/16/88に代表される山形系統とB/Victoria/2/87に代表されるVictoria系統がある。2008/09シーズンの国内分離株は1,915株で、内訳は山形系統25%、Victoria系統75%であり、後者が優位を占める両系統の混合流行であった。海外諸国においてもB型の流行株の90%はVictoria系統であった。これら分離株については9種類のフェレット参照抗血清を用いて抗原性解析が行われた。
抗原性解析:B型の中で主流を占めたVictoria系統分離株は、シーズン前半から本系統のワクチン株B/Malaysia/2506/2004からは抗原性が大きく変化しており、解析した分離株のほぼすべてがHI試験でB/Malaysia/2506/2004から8倍以上減少した変異株であった。諸外国でも同様の傾向が見られたことから、2009/10北半球シーズン向けのワクチン株は、変異株の代表であるB/Brisbane/60/2008類似株に変更された(4.ワクチン株の項参照)。
感染研で作製した孵化鶏卵馴化B/Brisbane/60/2008株に対するフェレット抗血清は、MDCK細胞分離株とほとんど反応しない。これは、すでに報告したように鶏卵への馴化によって糖鎖結合部位(197-199)に変異が入り、これによって糖鎖が欠失して抗原性が変化するためである。このため、感染研では、ワクチン株B/Brisbane/60/2008類似株に属し、MDCK細胞で分離されたB/Sakai(堺)/43/2008株に対する抗血清を作製して、これに対する反応性を基準にして、ワクチン株との抗原性の違いを評価した。その結果、シーズン後半には、ワクチン類似株が67%であった。一方、解析した中国株の大半は変異株で、これらはHA遺伝子系統樹上でも流行の主流とは異なる一群を形成していた。中国以外の諸外国では、南北アメリカでは71〜87%、南半球では56%がそれぞれB/Brisbane/60/2008類似株であった。
少数ながら国内で分離された山形系統株については、前シーズンのワクチン株B/Florida/4/2006抗血清とはほとんど反応せず、その類似株でHA遺伝子グループの異なるB/Bangladesh/3333/2007抗血清とよく反応した。諸外国で分離された山形系統株の大半は、依然B/Florida/4/2006類似株であった。
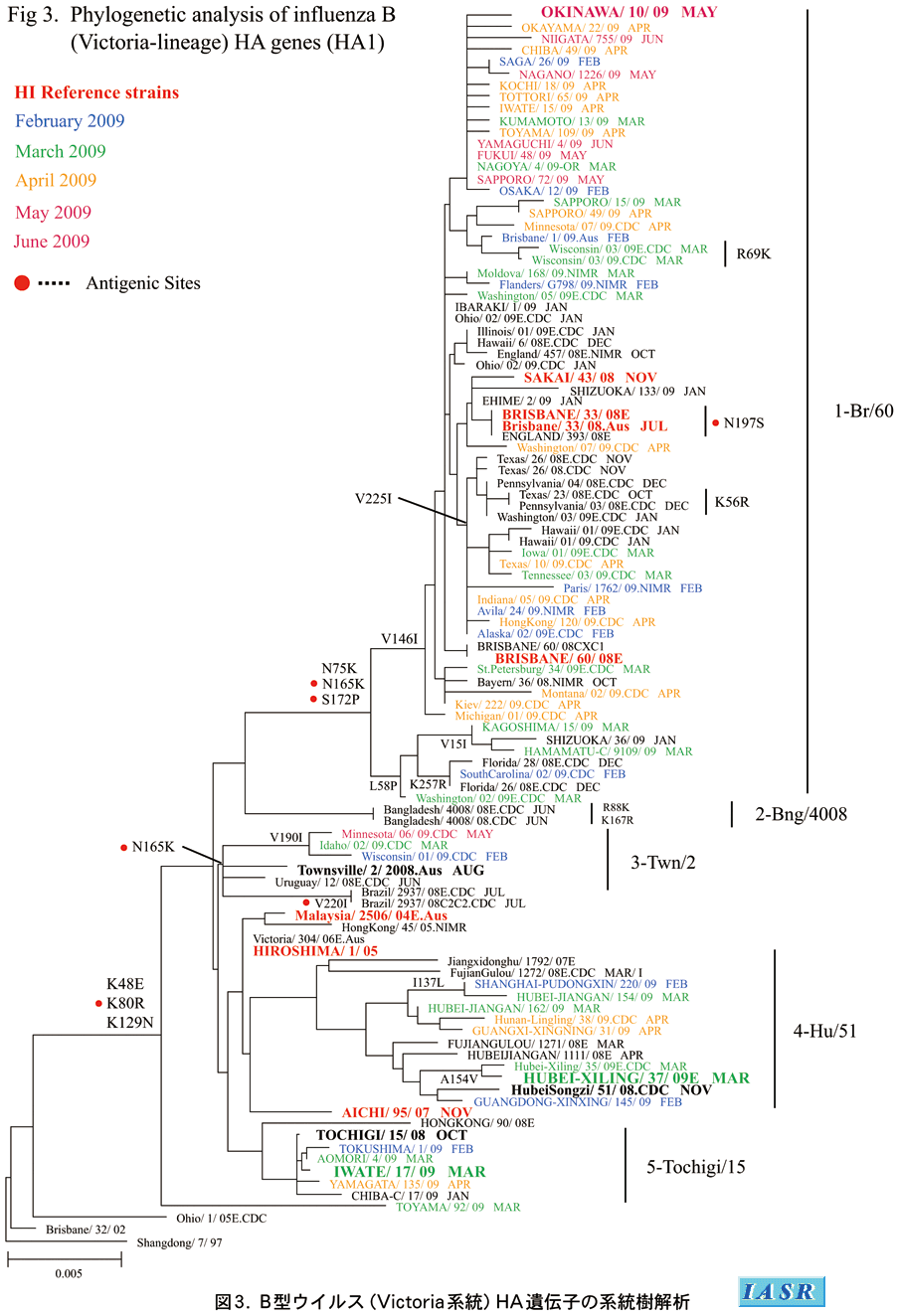
遺伝子系統樹解析:Victoria系統の系統樹解析(図3)では、シーズンを通じて流行株の大半はN75K、N165K、S172P置換をもちB/Brisbane/60/2008およびB/Sakai(堺)/43/2008で代表されるクレード1に属していた。また、B/Brisbane/60/2008とは抗原性の異なる多くの中国分離株は、B/Hubei-Xiling/37/2009で代表されるクレード4を形成していた。さらに、国内分離の一部の変異株はB/Iwate(岩手)/17/2009で代表されるクレード5を形成していた。
一方、山形系統株の系統樹は、B/Florida/4/2006で代表されるクレード1、B/Brisbane/3/2007で代表されるクレード2、B/Bangladesh/3333/2007で代表されるクレード3に大別されるが(図4)、国内外の流行株の大半はクレード3に属していた。
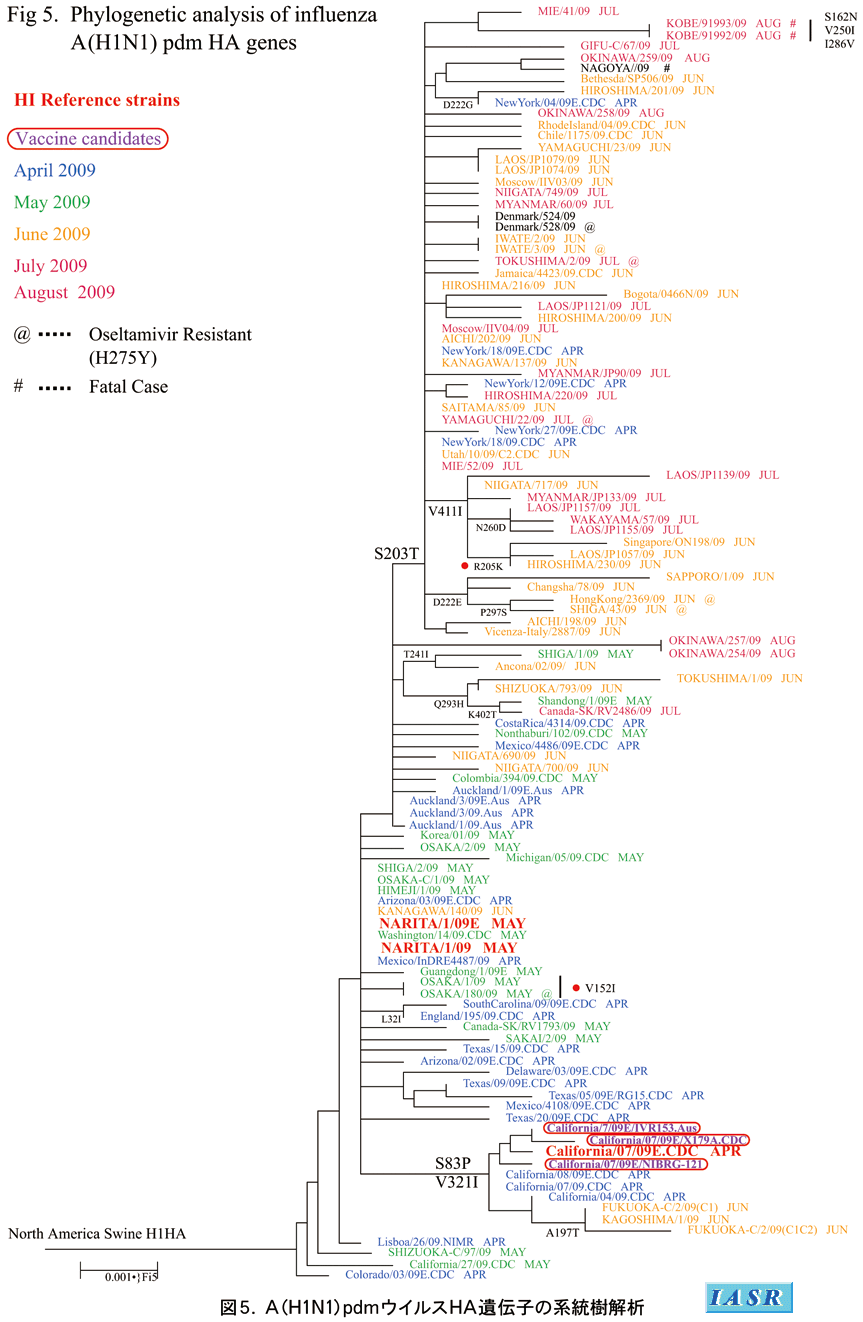
2-4)新型A(H1N1)pdmウイルス
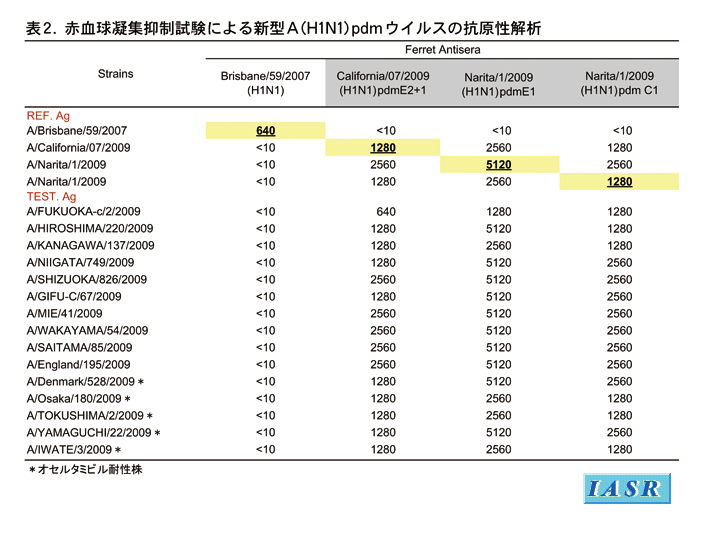
抗原性解析:国内では5月から流行が始まった新型A(H1N1)pdmウイルスは、第35週までに2,906株が分離され、第28週(7/6 〜7/12)以降は分離株の98%を占めた。国内分離株の137株について、国内初の分離株A/Narita(成田)/1/2009(孵化鶏卵およびMDCK細胞で分離したウイルス)、米国CDCから供与されたワクチン株A/California/7/2009、それぞれのウイルスに対するフェレット抗血清および七面鳥血球を用いてHI試験を実施した。また、季節性A/H1N1ウイルスとの抗原性を比較するためにA/Brisbane/59/2007抗血清との反応性も並行して評価した。
解析した国内分離株の99%はA/Narita(成田)/1/2009株およびA/California/7/2009株のホモ価と同じか2〜4倍反応性が高く、抗原性は均一であった。また、孵化鶏卵分離株とMDCK細胞分離株の間には抗原性の違いは見られなかった。このことは、新型A(H1N1)pdmウイルスは鶏卵馴化による抗原変異は起こっていないことを示している。また、散発的に分離された国内外のオセルタミビル耐性株も、重症死亡例から分離された株も、抗原的にはA/Narita(成田)/1/2009株またはA/California/7/2009株と区別されなかった。一方、新型A(H1N1)pdmウイルスは、季節性A/H1N1ウイルスA/Brisbane/59/2007に対する抗血清とは全く反応せず、抗原性が大きく異なっていた。現時点でA/California/7/2009抗血清のホモ価から4倍程度減少した国内分離株が数株見つかっているが、いずれもMDCK細胞での継代により起こった変異であり、原株での変異ではなかった(表2)。同様に諸外国における分離株もA/California/7/2009類似株がほとんどを占め、変異株が分離されているのは南半球における総解析株の8%という最近の報告のみである。従って、世界中の新型A(H1N1)pdmウイルスの抗原性は均一で、ワクチン株A/California/7/2009と類似していたといえる。
遺伝子系統樹解析:新型A(H1N1)pdmウイルスのHA遺伝子系統樹解析により、国内外で分離されたすべての株はA/Narita(成田)/1/2009株およびA/California/7/2009株を含む単一のクラスターに属し、遺伝的にも均一であることが示された(図5)。しかし、最近の国内株では、S203T置換を共通に持つサブクラスターを形成する傾向を示している。A/Osaka (大阪)/180/2009を除くオセルタミビル耐性株のすべては、この群に属していた。
3.抗インフルエンザ薬耐性株
3-1)A/H1N1ウイルス
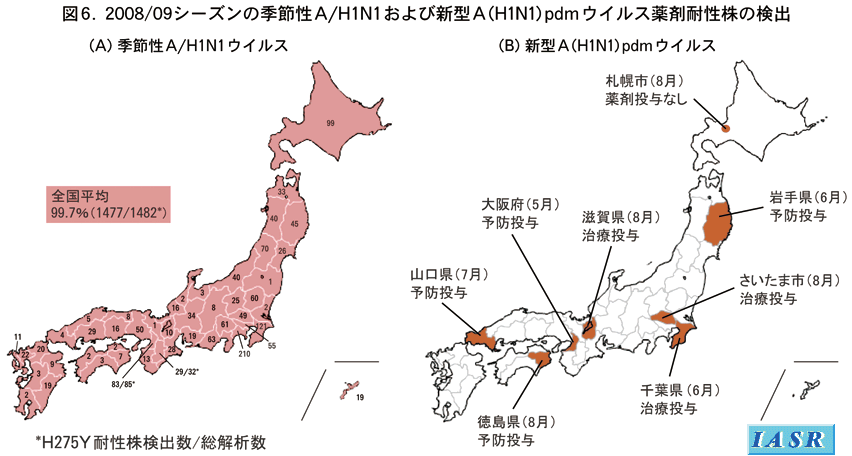
2007/08シーズンには、ヨーロッパ、北米などの海外諸国で、NA阻害剤オセルタミビルに対する耐性株が急速に広がり、大きな問題となったが、国内での発生は、数県に限られ、またその頻度は低く、解析したすべてのA/H1N1株の2.6%程度であった(IASR 29: 334-339, 2008)。一方、2008/09シーズンになると、わが国でも耐性株が急増し、全都道府県から検出報告があった。解析した1,482株中1,477株(99.7%)がNA蛋白に耐性マーカーH275Y置換を持っていた(図6A)。これら耐性マーカーを持つ株の5%に相当する分離株について、化学発光系を用いた薬剤感受性試験を実施したところ、ウイルスNA活性を半分に抑える薬剤濃度IC50値は、感受性株に比べて約300倍高く、酵素反応の上からも耐性であることが確認された。海外諸国においても同様で、ほとんどの国で分離されるA/H1N1ウイルスの95%以上はオセルタミビル耐性となっている。わが国の2008/09シーズンでは、季節性A/H1N1が全体の1/4以上を占めたことから、オセルタミビル耐性は医療現場に大きな影響を与えた。一方、オセルタミビル耐性株のすべては、別のNA阻害剤であるザナミビルに対しては、感受性を示していた。また、イオンチャンネルM2蛋白の阻害剤アマンタジンに対しては、遺伝子解析した株のうち、国内分離株は1/143(0.7%)、中国分離株は48/70(69%)に耐性株が検出された。
3-2)A/H3N2およびB型ウイルス
2008/09シーズンのA/H3N2およびB型分離株のうち、それぞれ172株、85株についてオセルタミビル、ザナミビル感受性試験を実施した。その結果、ザナミビルに対する感受性がやや低下した株がA/H3N2で6株見られたが、これら薬剤に対する明らかな耐性株は検出されなかった。海外においてもA/H3N2およびB型では耐性株は見つかっていない。一方、アマンタジンに対しては、諸外国と同様にA/H3N2の100%が耐性であった。
3-3)新型A(H1N1)pdmウイルス
国内外で分離されている新型A(H1N1)pdmウイルスは、すべてアマンタジンに対しては耐性である。新型ウイルスのNAとM遺伝子がユーラシア系統のブタウイルスに由来しており、この系統のウイルスのM遺伝子がアマンタジン耐性であることによる。一方、NA阻害剤については、わが国では2009(平成21)年10月下旬から全国組織でオセルタミビル耐性株サーベイランスが開始される。従って、現時点では臨床像から薬剤耐性が疑われた症例からの散発的分離例からで、5月以来8株の耐性株が見つかっている(図6B)。これらの大半は、感染者への濃厚接触後のオセルタミビル予防投与中または感染者の治療投与中に検出された。ただ、札幌市で分離された耐性株は、オセルタミビル未使用の患者から分離されたもので、耐性株によるヒト−ヒト感染が疑われた事例である。今のところ、これら耐性株による家族内および周囲への感染拡大は見られていない。また、これら耐性株はすべてザナミビルに対しては感受性であった。
世界では、欧州で4株、南北アメリカで16株、西太平洋地域で19株が検出されており(WHO, WER 84:453-459, 2009)、多くはオセルタミビル予防投与中または治療投与中における薬剤選択圧によるもので、散発的な耐性株の発生であった。
4.ワクチン株
2008/09シーズン前半には、国内外ともにA型、B型のいずれについても、2008年の南半球における流行株の抗原性、遺伝子系統がほとんど変化していなかった。従って、今冬(2009/10シーズン)の北半球向けの季節性ワクチン株としては、A/Brisbane/59/2007(H1N1)類似株、A/Brisbane/10/2007(H3N2)類似株(日本は製造株としてA/Uruguay/716/2007を採用)、B/Brisbane/60/2008(Victoria系統)類似株がWHOから推奨された。
一方、シーズン後半には、A/H3N2亜型ウイルスにおいて抗原性が大きく変化して、A/Perth/16/2009類似株が世界の主流となった。さらに、5月以降は新型A(H1N1)pdmウイルスによるパンデミックが起こり、分離株のほぼ大半を占めるようになった。季節性A/H1N1ウイルスはまだ消滅してはいないが、分離頻度は各国とも極めて低くなってきたことや、新型ウイルスに対する免疫の確保が優先されることから、WHOによる2010年の南半球向けワクチン株としては、A/H1N1亜型は季節性のソ連型から新型A(H1N1)pdm株へと置き換え、また、比較的大きな抗原変異が起こったA/H3N2ではワクチン株の変更がなされた。その結果、2010年の南半球向けのワクチン株としては、A/California/7/2009(H1N1)pdm類似株、A/Perth/16/2009(H3N2)類似株、B/Brisbane/60/2008(Victoria系統)類似株が推奨された(WHO, WER, 84, No.41, 421-432, 2009および本号17ページ)。
2008/09シーズンのワクチン株と流行株との一致性の評価
各年度のインフルエンザワクチン株は、上記のように国内外の流行株の分析による流行予測、前年度のワクチン接種前後のヒト血清中の抗体と流行株との反応性、流行前のワクチン株に対する国民の抗体保有調査の結果(http://idsc.nih.go.jp/yosoku/Flu/Serum-Flu2008.html)、次期ワクチン製造候補株の適性(孵化鶏卵での増殖効率および継代による抗原性、遺伝子の安定性など)などを総合的に評価して選定される。また、ワクチン株選定の時期は、2月にWHOによって決定される北半球向け推奨株を参考にして、3月末までに選定され、厚生労働省健康局長により5月〜6月までに正式に決定される。従って、ワクチン株の選定は、実際の流行が始まる9〜10カ月前までの成績に基づいて行われている。
株サーベイランスはWHOグローバル・サーベイランス・ネットワーク(GISN)により、地球規模で実施されるように改善されてきたため、流行予測精度が過去に比べて飛躍的に向上しているが、流行予測を早い時期に行わざるを得ないため、ワクチン株と流行株が結果的に一致しない場合がある。このような背景を踏まえて、前年度(2008/09シーズン)のワクチン株と実際の流行状況とを振り返って評価した。
2008/09シーズンに向けた本邦でのワクチン株は、A/Brisbane/59/2007(H1N1)、A/Uruguay/716/2006(H3N2)(A/Brisbane/10/2007類似株)、B/Florida/4/2006(山形系統)であり、上記の総合分析をもとに、2008年3月に選定され、6月に公表された(IASR 29: 307-309, 2008)。A/H1N1亜型ウイルスは流行最盛期(2008年12月下旬〜2009年2月下旬)で94%、流行終盤(3月以降)においても85%の流行株はワクチン株と抗原性が一致していた。A/H3N2亜型は最盛期でワクチン株との一致性は72%、終盤から第30週までの小規模流行では逆に75%がワクチン株から8倍以上の抗原変異を示していた。一方、B型のワクチン株は山形系統から選定されていたが、実際の流行株は、抗原性ではほとんど交叉反応しないVictoria系統であり、ワクチン株とは一致していなかった。ウイルス分離株の割合に基づいて算出した各ウイルスの流行比率は、新型A(H1N1)pdmウイルスが発生する以前においては、A/H1N1が47%、A/H3N2が26%、B型が27%であった。この結果は、ワクチン株の一致性と各ウイルスによる流行規模は必ずしも相関していないことを示している。
現行ワクチンの皮下接種は、ウイルス感染そのものを完全には阻止できないが、個人レベルでは、重症化、死亡のリスクを減らすことが示されている。インフルエンザの流行規模は免疫保有状況のみで規定されるわけではないが、2008/09シーズンの結果は、ワクチン株が一致していても、接種率が40%を下回る状況では、現行のワクチンは社会全体における流行動態には大きな影響を与えないことを示唆している。
本研究は「厚生労働省感染症発生動向調査に基づくインフルエンザサーベイランス」事業として全国76地研*との共同研究として行われた。また、ワクチン株選定にあたっては、ワクチン接種前後のヒト血清中の抗体と流行株との反応性の評価のために、特定医療法人原土井病院臨床研究部池松秀之部長、新潟大学大学院医歯学総合研究科国際感染医学講座公衆衛生学分野齋藤玲子博士、鈴木宏教授からの協力を得た。また、本研究には、島袋梢、松浦純子、望月菊各氏の協力を得た。海外からの情報はWHOインフルエンザ協力センター(米CDC、英国立医学研究所、豪WHO協力センター)から提供された。本稿に掲載した成績は全解析成績の中から抜粋したものであり、残りの成績は既にNESIDの病原体検出情報で各地研に還元された。また、本稿は上記研究事業の遂行にあたり、地方衛生研究所全国協議会と感染研との合意事項に基づく情報還元である。
国立感染症研究所
インフルエンザウイルス研究センター第1室・WHOインフルエンザ協力センター
小田切孝人 小渕正次 岸田典子 氏家 誠 徐 紅 高下恵美 伊東玲子 土井輝子
安楽 茜 江島美穂 菅原裕美 田代眞人
独立行政法人製品評価技術基盤機構 加藤裕美子 小口晃央 堀川博司 藤田信之
地方衛生研究所インフルエンザ株サーベイランスワーキンググループ*
 今月の表紙へ戻る
今月の表紙へ戻る
 IASRのホームページに戻る
IASRのホームページに戻る Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}